間隙水に棲む貝形虫類-垣間見る驚異的な種多様性- 2010.02.23
著者: 塚越 哲 (静岡大学理学部)
浜辺の砂を掘ると水が湧いてきます。この水を間隙水といいますが、この間隙水からは、これまで実に20以上の動物門の生物が世界中から報告されています。その中には、腹毛動物や動吻動物のように日常的にあまり知られていない生物から、刺胞動物(クラゲの仲間)、軟体動物(巻貝の仲間)それに私たちと同一祖先をもつ脊索動物(ホヤの仲間)までもが含まれています。間隙水の中は我々が日常目にすることのない場所ですが、多種多様な微小動物が独自の世界を作って生息しているのです。ここでは、微小甲殻類である貝形虫類(カイミジンコ、Ostracoda)を例にして、その驚異的な種数と生物多様性・生物進化研究への展望についてご紹介します。
間隙性生物と貝形虫類
貝形虫類(オストラコーダ/Ostracoda、和名ではカイミジンコとよばれる)とは節足動物・甲殻類に含まれる1分類群で(図1・2)、二枚貝のように体全体が左右2枚の殻(背甲)に包まれていることが特徴である。種多様性は極めて高く、現世の海や湖、河川などから8,000種余が知られており、化石種をも含めるとこれまでに30,000を優に越える種が知られている。生活様式は遊泳するもの、浮遊するものもあるが、多くのものは底生であり、砂の上や海藻などに生活している。
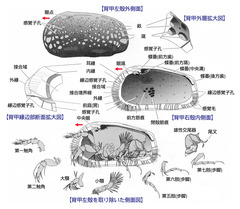
図1: 貝形虫類の一般的体制
節足動物・甲殻類である貝形虫類は、多くのものが1mm以下と小型ながら、機能分化した付属肢や感覚器官を備えた背甲など、複雑な形質を備える。間隙性種の場合は、各付属肢はより繊細であり、またしばしば第7肢を欠くものがある。(Horne 2002より改変引用)
(クリックで拡大します)
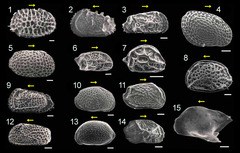
図2: 表在性貝形虫類の背甲の電子顕微鏡写真
日本周辺に生息する表在性の貝形虫類。背甲の外形、表面彫刻は種ごとに変化に富む。
1 Bradleya albatrossia, 2 Abrocythereis sp., 3 Cornucoquiba tosaensis, 4 Loxoconcha japonica, 5 Pistocythereis bradyi, 6 Semicytherura yajimae, 7 Hemicytherura sp., 8 Aurila spinifera 9 Trachileberis sp., 10 Aurila sp. A, 11 Aurila sp. B, 12 Yezocythere hayashii, 13 Cythere japonica, 14 Schizocythere kishiouyei, 15 Kobayashina hyalinosa。各スケールバーは100μm、黄色の矢印は各個体の前方を示す。
(クリックで拡大します)
ここでは底生貝形虫類の中で、特に間隙水中で生活するものについて解説する。間隙水とは、海や川などの水域において、堆積物の隙間を満たす水のことである。海水浴などで砂浜を掘ると水が染み出してくることは誰しも知ることであるが、この水が間隙水である。実際には海水浴に使われる浜辺の場合、砂の粒子が細かすぎて作られる間隙が小さいため、貝形虫類はあまり見られない。しかし、いわゆる「砂利浜」と呼ばれるような粗い砂や小石で構成される浜では間隙空間が大きくなるために、たくさんの生物の住処が提供されることとなる。そこには様々な生物が生息していることが知られている(図3・4)。

図3: 試料採集と間隙環境の概念図
A. 汀線付近における掘削。B. 間隙環境の概念図(Ward et al. 2000より改変引用)。
(クリックで拡大します)
間隙水中に棲む生物=間隙性生物の存在は、20世紀初頭に認識され、その後現在まで数々の生物が報告されてきた。現生する動物はおよそ35の門(Phylum)に分類されるが、その中の20以上の動物門に属する生物が、この間隙水中からも発見されているのである(伊藤1985)。つまり、節足動物や刺胞動物、軟体動物など、多くの動物がいわばミニチュアとなってこの間隙水中に適応し、一つの世界を作っている。
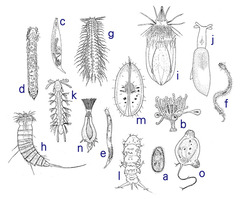
図4A: 様々な間隙性動物
a 繊毛虫類(原生生物)、b 刺胞動物、c 渦虫類(扁形動物)、d 二胚虫類(中生動物)、e 顎口動物、f 線虫類(線形動物)、g 腹毛動物、h 動吻動物、i 胴甲動物、j 軟体動物、k 多毛類(環形動物)、l クマムシ類(緩歩動物)、m 貝形虫類(節足動物)、n コケムシ類(外肛動物)、o 尾索類(脊索動物)。(伊藤1985より改変引用)
(クリックで拡大します)
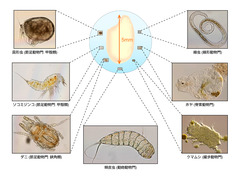
図4B: 様々な間隙性動物
米粒の大きさと比較した間隙性動物の光学顕微鏡写真。円の中にあるのが米粒の大きさと比較したもの。(資料提供: 静岡大学創造科学技術大学院・田中隼人)
(クリックで拡大します)
間隙水に適応した生物は、狭い空間で生活するためにおしなべて小型化している。貝形虫類の場合も、成体であっても体長0.2mmから0.4mm程度にとどまり、砂の表面や海藻に棲む表在性のものに比べて極めて小さい(図5)。体サイズが小型化するだけでなく、外部形態の単純化、目の退化、大卵少産、色素の消失などの共通した特徴を見ることができるが、これらは貝形虫類だけにとどまらず、間隙性生物一般に当てはまる事柄である。これは光がなく、また大型の捕食者が存在しえない空間で間隙性生物たちが独自の進化を遂げた結果であるといえる。しかし間隙性生物は、砂の隙間というヒトの視界から遮られた世界に棲む上、体サイズが一段と小さいことから、その実態をとらえた研究はあまりなされてこなかった。貝形虫類の場合には、種ごとの特徴が現れる背甲の形質にも乏しいため、表在性の種に比べてその分類学的多様性に関する情報も断片的なものにとどまっていた。
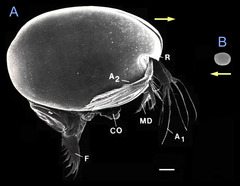
図5: 表在性種と間隙性種の大きさの比較
A 表在性種Vargula hilgendorfii (Vanner and Abe 1992より抜粋)。B Polycope japonica。V. hilgendorfiiはウミホタルとしても知られる。P. japonicaは、V. hilgendorfiiと同一の祖先を共有する仲間であるが、間隙生活に適応するために著しい矮小化がみられる。スケールバーは300μm、黄色の矢印は各個体の前方を示す。
(クリックで拡大します)
日本国内においても貝形虫類はもちろん、間隙性生物に関する一般的な理解は乏しいものであるが、唯一1985年に故伊藤立則博士によって著された「砂の隙間の生き物たち」は、間隙性生物全般について解説された極めて優れた書である。この書は既に絶版となってしまったが、筆者自身も大学院生時代にこの書に出会い、間隙性生物が織り成す世界の多様さに感銘を受け、現在の研究に着手する動機付けとなった。
日本における間隙性貝形虫類の多様性
筆者が間隙性貝形虫類の研究に着手したのは2000年のことであるが、この時点で日本における間隙性貝形虫類はわずか5種しか知られていなかった(Schornikov 1975; Hiruta 1983, 1989, 1991)。しかしその後、学生たちと共に駿河湾~相模湾の海岸線約250km、約40地点を調査した結果、この地域だけで現在までに約50種を確認し、それらのほぼすべてが未記載種であった(図6)。日本の貝形虫類の研究は、1910年代に始まり、現在までに現生種約350種が知られている(Ikeya et al. 2003)。この数字は日本とその周辺海域すべてについて、90年余をかけて分類・記載が行われてきた結果得られた数字である。これに対し、駿河湾~相模湾沿岸の間隙性貝形虫類の種数は、限られた地域において限られた年数で行われた調査によって得られた種数である。
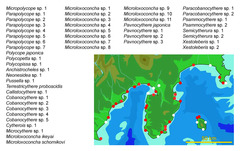
図6: 駿河湾~相模湾沿岸における間隙性貝形虫類の種多様性
2009年現在48種が確認されているが、既知種は5種のみであり、残りは全て未記載種である。丸印は試料採集地点で、赤丸は間隙性貝形虫類が産出した地点。(データは、Watanabe et al. 2008、静岡大学理学部地球科学科卒評論文・渡辺 聡2002MS、同・東 亮一2005MS、同・高橋英明2006MS、同・田中隼人2007MS、同・高橋一真2008MS他より引用)
(クリックで拡大します)
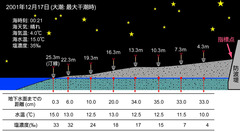
図7: 下田市大浦における試料採集の概念図
スコップの上に記されている数値は、防波堤上の指標点からの距離。大潮の最大干潮時に採集するため、春から夏にかけては昼間、秋から冬にかけては夜間に採集を行なう。(静岡大学理学部地球科学科卒業論文・渡辺 聡2002MSより改変引用)
(クリックで拡大します)
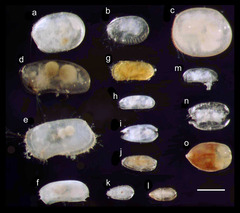
図8: 沖縄本島・瀬底より産出した間隙性貝形虫類の光学顕微鏡写真
産出した全ての種が未記載種である。a Parapolycope sp.、b Polycopiella sp.、c Metapolycope sp.、d Anchistrocheles sp.、e Orlovibairdia sp.、f Pussella sp.、g Callistocythere sp.、h Cytheroid sp.、i Microcythere sp.、j Cobanocythere sp. 1、k Cobanocythere sp. 2、l Parvocythere sp.、m Microloxoconcha sp.、n Cobanocythere? sp.、Xestoleberis sp. i, k, l, n, oは背面視、他は左側面視。スケールバーは200μm、左が前方。
(クリックで拡大します)
もし日本全体の海域で間隙性貝形虫類の詳しい調査を行うことをすれば、その種数はこれまで私たちが認識してきた貝形虫類の種多様性に関する概念を根本的に塗り替えるものとなるかもしれない。また、この種多様性はより南方において高いことを伺わせる結果も得られている。例えば、下田市大浦における定点観測では、汀線に垂直方向に観測点をとり(図7)、2カ月おきに2年間(計7回)試料採集を試みた結果、この地点からは7種の間隙性貝形虫類が確認された。これに対し、沖縄県本部町瀬底において同様の方法で2回の試料採集を行った結果、既に15種以上の貝形虫類が確認された(図8)。ここで確認された種も、そのほとんどすべてが未記載種であった。現在、沖縄のサンゴ礁に生息する貝形虫について、その多様性の実態を明らかにするべく調査を再開している。狭い海域に限っても、駿河湾~相模湾沿岸以上の高い種多様性が観測できるかもしれない。
分類学的多様性把握の鍵・種分類
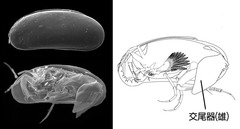
図9: 間隙性貝形虫類Microloxoconcha属の体制
体サイズは小さいが、間隙性貝形虫類の雄は発達した交尾器をもつ。左上段は左側面より、同下段は背甲の左殻を取り除いたもの。右はその体制の概念図。(資料提供: 静岡大学創造科学技術大学院・東 亮一)
(クリックで拡大します)
分類学的多様性を議論するには、基本単位である「種」レベルでの信頼性の高い分類が不可欠である。分類基準が曖昧なままでは、種多様性の評価を行うことはできない。貝形虫類は、体全体を背甲が覆うという特殊な体制をもち、海生種ではそのほとんどが雌雄異体で、交尾によって生殖を行う。したがって、雄は雌の体を包む背甲をこじ開け、その内部に位置するメスの交尾器に精子を送るべく交尾をする必要がある。そのため雄は体全体の1/4から1/3に達するほど発達した交尾器を尾部に備える場合が多い(図1・9)。またその多様な形態は、雌に対する接触刺激を作り出すことによって、雌は同種の雄か否かを区別する、すなわち生殖的隔離に寄与していると考えられる(Tsukagoshi 1988)。
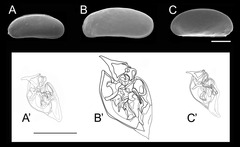
図10: Microloxoconcha属3種の背甲形態と交尾器形態の比較
背甲(上段の電子顕微鏡写真、スケールバーは100μm)ではサイズ以外には種ごとの違いを見出すことは難しいが、交尾器(下段のスケッチ、スケールバーは50μm)の形態は種ごとに明確な差が現れる。A, A' Microloxoconcha ikeyai, B, B' M. sp. 1, C, C' M. sp. 2。(AはWatanabe et al. 2008、B、CはHigashi and Tsukagoshi 2008より改変引用)
(クリックで拡大します)
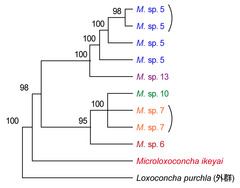
図11: DNA塩基配列による系統推定
COⅠ領域の塩基配列の比較からベイズ法によって得られた分岐図。数値はベイズ信頼度を表す。同一地点から得られた試料は半カッコでくくってある。(資料提供: 静岡大学創造科学技術大学院・東 亮一)
(クリックで拡大します)
従って貝形虫類では、この雄の交尾器の形態を比較することによって、種の違いを正確にとらえることができる(図10)。交尾器で認識された形態群(この場合独立した種と考えられる)は、DNAの塩基配列の比較からも支持されており、1つのクラスターを形成することが確認できる(図11)。また、DNAの塩基配列の比較は、種間関係を推測する有力な手段にも用いられ、多様化した間隙性種の種分化の順序や分岐時間に関する考察の手助けにもなっている(図11)。
なぜ分類学的多様性が高いのか
これまで研究された地域における間隙性貝形虫類の高い種多様性は、その地域にだけの現象ではないかもしれない。もしそうであるなら、間隙性貝形虫類の実際の種数は、表在生種のそれを凌駕する可能性さえある。同時に、間隙水中に生息する20門にも及ぶ動物群に同様な高い種多様性が当てはまるとしたら、海洋無脊椎動物全体の分類学的多様性の概念そのものをも大きく塗り替えることになるであろう。高い種多様性の理由は、なんであろうか? それは砂間間隙という、閉鎖的な生活場所にその一因があるのかもしれない。貝形虫類は浮遊幼生期をもたないため、比較的分散能力に乏しいと考えられている。表在生種の場合、卵や幼体ばかりでなく成体も流れ藻や表層堆積物の移動によって遠方へ移動すると考えられるが、堆積物の隙間に潜り込んで生活する間隙性種は、そのような移動手段をもたないのかもしれない。また、堆積物中の間隙空間は、実際には私たちが予想する以上の様々な微環境を作り出しているとも考えられる。体サイズそのものが非常に小さい間隙性種は、そこに成立する様々な微環境の違いに対して個々に適応して生活しており、これが高い種多様性を支えているという可能性も想定される。
間隙性貝形虫類に関する研究の今後
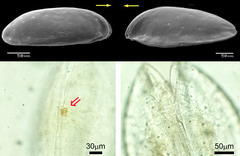
図12: 眼の退化
間隙性貝形虫類Cobanocythere属2種を比較すると、左の種は明確な眼を持つ(赤い矢印)が、右の種は完全に消失している。上段は背甲を側面から見た電子顕微鏡写真(黄色い矢印は体の前方を示す)。下段は背甲を上方から光学顕微鏡で観察したもの。(資料提供: 静岡大学大学院理学研究科・高橋香里)
(クリックで拡大します)
間隙性貝形虫類に見られた種多様性の高さは、筆者自身の予想をはるかに上回るものであったが、その実態の把握には記載的研究を継続する必要があり、これ以外に解決の方法はありえない。このためには長い時間をかけた研究の継続が不可欠である。こうした種多様性への興味とは別に、間隙性貝形虫類には生物進化学的にも注目すべき点が多くあり、今後の研究課題となっている。
一つは、眼の退化に関する問題である。洞窟や深海のような暗黒環境では、多くの生物が眼を退化させている。間隙環境もまた、光の届かない暗黒環境であるが、その度合は堆積物の深さによって異なる。例えば砂底表面から数10cmの深さは暗黒環境といってよいが、1cm程度の深さならば、波浪などによって簡単に白日の下にさらされることになる。事実、間隙性貝形虫では同じ系統(同属)の種同士であっても、眼を持つ種と退化している種があることが明らかになってきた(図12)。これは動物の眼の退化という普遍的な問題に対して、間隙性生物を用いることによって、これまでにないアプローチが可能となる(Kaji and Tsukagoshi, 2010)。
さらに間隙性貝形虫類の中には、「生きている化石」とよぶにふさわしい、原始的な体制を保持したまま現在まで生き延びている分類群も多数確認されている。間隙空間が外界から遮断され、大型の捕食者が入り込めない空間を形成しているとすれば、そのような分類群が貝形虫類に限らず多数存在していることも考えられる。また、深海に生息する分類群とのつながりも示唆する事例も見られる。例えば、非常に近縁な2つの種(同属種)が、それぞれ沖縄のサンゴ礁海岸の深さ10cm程度の砂間間隙と、水深1,000mを越える北極海の海底とから産出している例もある(塚越, 2004)。
間隙空間を生息場とする生物たちに関する研究は、生物の種多様性に関する概念を大きく塗り替える可能性をもつと共に、生物進化について新たな視点を与える可能性をも秘めている。
参考図書
- 1) 池谷仙之・阿部勝巳(著) 太古の海の記憶 オストラコーダの自然史. 東京大学出版会 東京.
- 2) 岩槻邦男・馬渡峻輔(編) 生物の種多様性 バイオディバーシティーシリーズ1. 裳華房. 東京.
- 3) 片倉晴雄・馬渡峻輔(編) 動物の多様性 シリーズ21世紀の動物科学2. 培風館. 東京.
- 4) 池谷仙之・棚部一成(編) 古生物の生活史 古生物の科学3. 朝倉書店. 東京.
- 5) 瀬戸口烈司・小沢智生・速水 格(編) 古生物の進化 古生物の科学4. 朝倉書店. 東京.
引用文献
- Ikeya N, Tanaka G, Tsukagoshi A (2003) The database of Japanese fossil type specimens described during the 20th Century (Ostracoda). Pp. 37-131. In: Ikeya N, Hirano H, Ogasawara K (Eds) The database of Japanese fossil type specimens described during the 20th Century (Part 3), Palaeontological Society of Japan (Special Papers 41), Tokyo.
- 伊藤立則 (1985) 砂の隙間の生き物たち. 241p., 海鳴社.
- Higashi R, Tsukagoshi A (2008) Two new species of Microloxoconcha (Crustacea: Ostracoda: Podocopida) from Japan, with a redescription of the genus. Species Diversity 13: 157-173.
- Hiruta S (1983) A new species of the genus Polycope Sars from the Inland Sea of Japan (Ostracoda: Cladocopina). Proceedings of the Japanese Society of Systematic Zoology 26: 1-10.
- Hiruta S (1989) A new species of marine interstitial Ostracoda of the genus Microloxoconcha Hartmann from Hokkaido, Japan. The Proceedings of the Japanese Society of Systematic Zoology 39: 29-36.
- Hiruta S (1991) A new species of marine interstitial Ostracoda of the genus Psammocythere Klie from Hokkaido, Japan. Zoological Science 8: 113-120.
- Horne DJ, Cohen A, Martens K (2002) Taxonomy, morphology and biology of Quaternary and living Ostracoda. Pp. 5-36. In: Holmes JA, Chivas AR (Eds) The Ostracoda -Application in Quaternary Research. American Geophysical Union (Geophysical Monograph 131), Washington, DC.
- Kaji T,Tsukagoshi A. (2010). Heterochrony and modularity in the degeneration of maxillopodan nauplius eyes. Biological Journal of the Linnean Society 99: 521-529.
- Tsukagoshi A (1988) Reproductive character displacement in the ostracod genus Cythere. Journal of Crustacean Biology 8: 563-575.
- 塚越 哲 (2004) 種多様性研究と古生物学: 間隙性貝形虫類を例として。化石 75: 18-23.
- Schornikov EI (1975) Ostracod fauna of the intertidal zone in the vicinity of the Seto Marine Biological Laboratory. Publications of the Seto Marine Biological Laboratory 22: 1-30.
- Ward JV, Malard F, Stanford JA, Gonser T (2000) Interstitial aquatic fauna of shallow unconsolidated sediments, particularly hyporheic biotopes. Pp. 41-58. In: Wilkens, H., Culver, D. C. and Humphreys, W. F. (Eds) Subterranean Ecosystems. Elsevier, Amsterdam.
- Watanabe S, Tsukagoshi A, Higashi R (2008) Taxonomy and ecology of two new interstitial Ostracoda (Podocopida: Cytheroidea) from Shimoda, central Japan. Species Diversity 13: 53-71.
東京都目黒区上目黒1丁目26番1号
中目黒アトラスタワー313
- TEL
- 03-3713-5635
当財団は、ナチュラルヒストリーの研究の振興に寄与することを目的に、1980年に設立され、2012年に公益財団法人に移行しました。財団の基金は故藤原基男氏が遺贈された浄財に基づいています。氏は生前、活発に企業活動を営みながら、自然界における生物の営みにも多大の関心をもち続け、ナチュラルヒストリーに関する学術研究の振興を通じて社会に貢献することを期待されました。設立以後の本財団は、一貫して、高等学校における実験を通じての学習を支援し、また、ナチュラルヒストリーの学術研究に助成を続けてきました。2024年3月までに、学術研究助成883件、高等学校への助成127件を実施しました。