シオガマギクの系統地理 2009.08.31
著者: 藤井 紀行 (熊本大学理学部)
日本列島には今現在、約5,000~6,000種もの植物(維管束植物以上)が知られています。その中の約40%は日本に固有な植物と言われており、このことは日本列島の中において活発な種分化が生じたことを物語っています。日本列島はアジア大陸の東端に位置し、そこを舞台にさまざまな生物が行き交い、または新種が生まれてきたものと想像されます。今日本列島に見られる植物はいったいどこからやってきたのでしょうか。またどのように生じてきたのでしょう。私はそうした疑問に答えるために、DNAを使った系統地理学的な研究を進めてきました。
対象種シオガマギクとは

図1: シオガマギクの形態変異
左上: 長野県大鹿村三伏峠(トモエシオガマ)
右上: 愛知県豊橋市(ミカワシオガマ)、後ろに見えるのはシラタマホシクサ
左下: 韓国ソラク山、ここの集団は花冠が白い個体ばかりであった。
下中段: サハリン(ビロードシオガマ)、植物体に毛が多く生えており、白っぽくみえる
右下: 山梨県乙女高原(いわゆるシオガマギク)
(クリックで拡大します)
ここでは研究の対象種としてシオガマギク(Pedicularis resupinata L.)という植物を用いました(図1)。ゴマノハグサ科の多年生草本で、少々湿った場所を好む植物です。東アジアに広く分布しており、日本でも九州から北海道まで広く分布しています。シオガマギクの興味深い点としては地域的に形態変異が大きいことが挙げられます。本州中部の亜高山帯の集団はトモエシオガマ、愛知県の三河地方や岐阜県南部の低湿地集団はミカワシオガマと種内分類群として区別されることがあります。これらの地域固有の変異は、日本列島内の地域的な環境の違いや地史的な変遷の歴史の中で生じてきたものと想定されますが、具体的にどのようにして分化してきたのでしょうか。そこで私は各地の地域固有のシオガマギク類を含めてサンプリングを行い、集団間の遺伝的な関係を調べてみることにしました。
葉緑体DNAの地理変異
まずは分布域をできるだけ網羅するように、シオガマギクを採集しました。合計48集団172個体を解析に用いました。その中には朝鮮半島から6集団、サハリンから6集団、千島列島から4集団の海外の集団を含めました。また形態的にトモエシオガマと同定された2集団(蝶ヶ岳、三伏峠(長野県))、ミカワシオガマと同定された2集団(石巻山(愛知県)、保古の湖(岐阜県))も同時に解析に加えました。採集してきた葉っぱを実験室でDNA抽出を行い、遺伝子増幅法(PCR法)を用いて、特定の遺伝子領域(葉緑体DNAのtrnL~F領域)を増幅し、塩基配列を決定しました。そのデータをきれいに整列することによって塩基置換や挿入・欠失といった種内多型を検出することができました。その結果から、各個体のDNAタイピングを行いました。
9つのDNAハプロタイプ
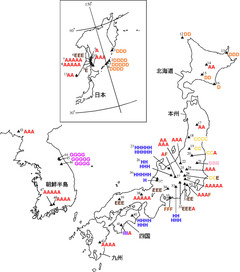
図2: シオガマギクのDNAタイプの地理的変異
葉緑体DNAのtrnL-Fの遺伝子間領域の塩基配列データによるDNAタイピングの結果、9種類のタイプが検出された。Aタイプが広域分布している。集団名はFujii (2003)を参照のこと。
(クリックで拡大します)
全ての個体を解析した結果、合計9つのDNAハプロタイプを検出することができました。それらにタイプA~Iと名付けて区別し、地図上にその結果を載せたものが図2になります。最も普遍的に見られるAタイプは、千島列島では見られませんでしたが、それ以外の地域ではまんべんなく分布していることが分かります。それ以外のタイプはある程度地理的なまとまりを持ちながら分布しています。例えば、Dタイプは北海道と千島列島のみに分布し、Cタイプは東北南部から関東にかけてのみ、Hタイプは本州中部から四国にかけてのみ、Gタイプは朝鮮半島中部のみといった感じです。BタイプやIタイプは特定の集団のみに見られる固有なタイプでした。
大きな2つの系統の存在!
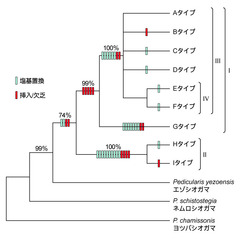
図3: シオガマギクのDNAタイプ間の系統樹
葉緑体DNAのtrnL-Fの遺伝子間領域の塩基配列データによるシオガマギクのDNAタイプ間の最節約系統樹。枝の上の数字は各枝の統計的な支持率を示す(ブートストラップ確率)(Fujii 2003を改変)。
(クリックで拡大します)
続いてタイプA~Iの間の系統関係を推定しました。この解析によりタイプ間の遺伝的な類縁性を知ることができます。最節約法を用いて推定した系統樹が図3です。解析の結果、大きな2つの系統が存在することが分かりました(系統I, II)。両者の統計的な支持率はほぼ100%なので、明瞭な2つの系統が日本列島の中に存在していることになります。両タイプの分布をもう少し詳しく見てみましょう。系統Iは広域に分布するタイプAを含み、今回解析した集団の分布域全体に広がっています。一方系統IIは、本州中部と四国にのみ分布が限定されています(H, Iタイプ)。
広域分布する系統I
系統Iを詳しく見ると、その中に系統IIIやIVといったまとまりがあることが分かります。さらに系統IIIとは異なる系統としてGタイプの存在が目につきます。Gタイプは朝鮮半島中部のソラク山やオデ山の集団です。図3を見ても分かるように同じ系統Iとは言っても系統IIIのタイプとは遺伝的なかなり異なっていることが分かります。朝鮮半島の北部と南部には広域分布のAタイプが見られますので、ソラク山やオデ山の地域の集団は何らかの理由により長期間隔離されていた可能性があります。
A~Fタイプが系統IIIを形成していますが、その中の遺伝的な分化は比較的小さいと言えます(図3)。しかしそれらの分布域は今回解析したほぼすべての地域を網羅しています。この結果は、系統IIIのシオガマギクが最近になって急速に分布域を拡大し、北方地域でDタイプ、東北地域でCタイプというように地域分化を起こしたことを示唆しています。
日本固有の系統IIの起源
日本の本州中部と四国のみに固有な系統IIはどのように生じてきたのでしょうか。系統IとIIはもともと共通な一つの系統だったはずです。それらが何らかの要因により2つの系統に別れたのです。この要因にはいくつかの可能性を考えることができますが、最も単純なストーリーは、以下のようなものだと思われます。系統IとIIの共通祖先の集団が日本列島に侵入した後、気候変動により一部の集団が日本列島に遺存的に残り、それが系統IIへと分化していった、という仮説です。つまり系統IIは大陸から移動してきたのではなく、日本列島の中で起源した系統なのではないかということです。
トモエシオガマとミカワシオガマの起源
さて、上で述べたトモエシオガマとミカワシオガマに着目してみたいと思います。トモエシオガマは図2のNo. 26と33の2集団になります。両集団とも系統IIのHタイプのDNAを持っていました。トモエシオガマでない集団にもHタイプは見られましたが、この結果はトモエシオガマが日本に固有な系統IIの一部から生じてきたことを示唆しています。一方、ミカワシオガマは図2のNo. 34と35の2集団になります。各々、EタイプとFタイプのDNAを持っていました。ここでも両タイプはミカワシオガマ固有のタイプではありませんでした。FタイプはNo. 35の集団固有のタイプですが、Eタイプはよく見るとサハリンからも検出されています(図2)。しかし系統解析の結果、両タイプは一つの系統を形成し(系統IV)、共通の祖先から分化していきたことが分かります(図3)。ミカワシオガマは広域分布する系統Iの中の一系統IVから分化してきたグループなのかもしれません。
最後に
シオガマギクは実際には東アジア(シベリア、モンゴル、満州などを含む)に広く分布しています。今回は財団の助成金により朝鮮半島のサンプルを加えることができましたが、シベリア、モンゴル、満州などのサンプルを加えるとまた解釈は変わってくる可能性があります。今後より厳密な解釈を行うためには大陸産のサンプルを増やして解析する必要があるでしょう。今回の研究の詳細は参考文献のFujii (2003)を参照してください。
参考文献
- Fujii N (2003) Chloroplast DNA phylogeography of Pedicularis resupinata (Scrophulariaceae) in Japan. Acta Phytotaxonomica et Geobotanica 54: 163-175.
- 山崎 敬 (1981) ゴマノハグサ科. 日本の野生植物 草本III合弁花類(佐竹義輔・大井次三郎・北村四郎・亘理俊次・冨成忠夫編). 平凡社. 東京.
- ジョン・C・エイビス (2008) 生物系統地理学 種の進化を探る(西田 睦・武藤文人監訳). 東京大学出版会. 東京.
東京都目黒区上目黒1丁目26番1号
中目黒アトラスタワー313
- TEL
- 03-3713-5635
当財団は、ナチュラルヒストリーの研究の振興に寄与することを目的に、1980年に設立され、2012年に公益財団法人に移行しました。財団の基金は故藤原基男氏が遺贈された浄財に基づいています。氏は生前、活発に企業活動を営みながら、自然界における生物の営みにも多大の関心をもち続け、ナチュラルヒストリーに関する学術研究の振興を通じて社会に貢献することを期待されました。設立以後の本財団は、一貫して、高等学校における実験を通じての学習を支援し、また、ナチュラルヒストリーの学術研究に助成を続けてきました。2024年3月までに、学術研究助成883件、高等学校への助成127件を実施しました。